学習成果
- 葉緑体および中心液胞を含む植物細胞にのみ存在するキーオルガネラを特定
- 中心体およびリソソームを含む動物細胞にのみ存在するキーオルガネラを特定
この時点で、真核細胞はより複雑な構造を持っていることは明らかであるべきである。原核細胞。 オルガネラは、同時に細胞内で様々な機能が起こることを可能にする。, それらの基本的な類似点にもかかわらず、動物細胞と植物細胞の間にはいくつかの顕著な違いがあります(図1参照)。
動物細胞は中心体(または中心小体のペア)およびリソソームを有するが、植物細胞は有さない。 植物細胞は、貯蔵に使用される細胞壁、葉緑体、原形質連絡体、および色素体、および大きな中央液胞を有するが、動物細胞は有さない。
練習問題

図1. (a)典型的な動物細胞および(b)典型的な植物細胞。,
植物細胞には動物細胞にはないどのような構造がありますか? 動物細胞には植物細胞にはないどのような構造がありますか?
植物細胞
細胞壁
図1bでは、植物細胞の図では、細胞壁と呼ばれる原形質膜の外側の構造が見られます。, 細胞壁は、細胞を保護し、構造的支持を提供し、細胞に形状を与える剛性被覆である。 真菌細胞およびいくつかの原生細胞はまた、細胞壁を有する。
原核細胞壁の主要成分はペプチドグリカンであるが、植物細胞壁の主要な有機分子はセルロースである(図2)。 栄養情報が食物繊維を指すとき、それは食品のセルロース含有量を指しています。

図2., セルロースは、1-4結合によって接続されたβ-グルコース分子の長鎖である。 図の各端にある破線は、一連のさらに多くのグルコース単位を示しています。 ページのサイズは、全体のセルロース分子を描くことが不可能になります。
葉緑体

図3. 葉緑体のこの簡略化された図は、外膜、内膜、チラコイド、グラナ、および間質を示しています。
ミトコンドリアと同様に、葉緑体も独自のDNAとリボソームを持っています。, 葉緑体は光合成において機能し、植物および藻類などの光独立栄養性真核細胞に見出すことができる。 光合成では、二酸化炭素、水、および光エネルギーは、グルコースと酸素を作るために使用されます。 これは植物と動物の大きな違いです:植物(独立栄養生物)はグルコースのような自分の食物を作ることができますが、動物(従属栄養生物)は有機化合物や食,
ミトコンドリアのように、葉緑体は外膜と内膜を持っていますが、葉緑体の内膜に囲まれた空間の中には、チラコイドと呼ばれる流体で満たされた膜嚢が相互接続されて積み重ねられています(図3)。 チラコイドの各スタックはgranum(複数形=grana)と呼ばれています。 内部の膜によって囲まれ、グラナを囲む液体は間質と呼ばれます。
葉緑体にはクロロフィルと呼ばれる緑色の色素が含まれており、光合成のために太陽光のエネルギーを取り込んでいます。 植物細胞と同様に、光合成原生生物も葉緑体を持っています。, いくつかの細菌も光合成を行いますが、葉緑体はありません。 それらの光合成色素は、細胞内のチラコイド膜に位置する。
Endosymbiosis
ミトコンドリアと葉緑体の両方にDNAとリボソームが含まれていることに言及しています。 なぜ疑問に思いましたか? 強力な証拠は、説明としてendosymbiosisを指しています。
共生は、二つの別々の種からの生物が密接に関連して住んでおり、典型的にはお互いに特定の適応を示す関係である。, エンドシンビオシス(endo-=within)は、ある生物が他の生物の中に住んでいる関係です。 内共生関係は自然に富む。 ビタミンKを産生する微生物は、人間の腸の中に住んでいます。 この関係は、ビタミンKを合成することができないため、私たちにとって有益であり、他の生物から保護され、大腸内に住むことによって安定した生息地と豊富な食物を提供されるため、微生物にとっても有益である。
科学者たちは、細菌、ミトコンドリア、葉緑体の大きさが似ていることに長い間気づいてきました。, また、ミトコンドリアと葉緑体には細菌と同じようにDNAとリボソームがあることも知っています。 科学者たちは、宿主細胞が好気性細菌およびシアノバクテリアを摂取したが、それらを破壊しなかったときに宿主細胞および細菌が相互に有益な内共生的関係を形成したと信じている。 進化を通じて、これらの摂取された細菌は、好気性細菌がミトコンドリアになり、光合成細菌が葉緑体になると、その機能に特化したようになりました。,
それを試してみてください
中央液胞
以前、我々は植物細胞の必須成分として液胞を言及しました。 図1bを見ると、植物細胞はそれぞれ、細胞の大部分を占める大きな中央の液胞を持っていることがわかります。 中央液胞は、環境条件の変化における細胞の水の濃度を調節する上で重要な役割を果たしています。 植物細胞では、中央液胞内の液体は、細胞内の流体によって引き起こされる外向きの圧力である膨圧を提供する。, あなたは数日間植物に水を注ぐことを忘れると、それが枯れることに気づいたことがありますか? それは、土壌中の水濃度が植物中の水濃度よりも低くなるにつれて、水は中央の液胞および細胞質から出て土壌中に移動するからである。 中央液胞が収縮するにつれて、細胞壁は支持されていないままになります。 植物の細胞壁への支持のこの損失は、しおれた外観をもたらす。 中心液胞が水で満たされると、それは植物細胞が拡大するための低エネルギー手段を提供する(実際にサイズが増加するためにエネルギーを費やすの, さらに、この液体は含んでいる無駄の苦い好みが昆虫および動物によって消費を落胆させるので草食を躊躇できます。 中央液胞はまた、種子細胞の発生中にタンパク質を貯蔵するために機能する。
動物細胞
リソソーム

図4. マクロファージは潜在的に病原性のある細菌を小胞に貪食し、それが細胞内のリソソームと融合して病原体を破壊することができる。 他の細胞小器官は細胞内に存在するが、簡単にするために、示されていない。,
動物細胞では、リソソームは細胞の”ごみ処理”です。”リソソーム内の消化酵素は、タンパク質、多糖類、脂質、核酸、さらには使い古したオルガネラの分解を助けます。 単細胞真核生物では、リソソームは、彼らが摂取する食品の消化とオルガネラのリサイクルのために重要です。 これらの酵素は、細胞質に位置するものよりもはるかに低いpH(より酸性)で活性である。, 細胞質内で起こる多くの反応は、低いpHでは起こらなかったので、真核細胞を細胞小器官に区画化する利点は明らかである。
リソソームはまた、細胞に入る可能性のある病気を引き起こす生物を破壊するためにそれらの加水分解酵素を使用する。 これの良い例は、あなたの体の免疫システムの一部であるマクロファージと呼ばれる白血球のグループで発生します。 食作用として知られているプロセスでは、大食細胞の原形質膜のセクションはinvaginates(折目)病原体を巻き込みます。, 病原体が内部にある陥入した部分は、その後原形質膜から自分自身を挟み込み、小胞になる。 小胞はリソソームと融合する。 その後、リソソームの加水分解酵素が病原体を破壊します(図4)。
動物細胞の細胞外マトリックス

図5. 細胞外マトリックスは、細胞によって分泌される物質のネットワークからなる。
ほとんどの動物細胞は、細胞外空間に物質を放出する。 これらの材料の第一次部品は糖蛋白質および蛋白質のコラーゲンです。, これらの物質を総称して細胞外マトリックスと呼びます(図5)。 細胞外マトリックスは、組織を形成するために細胞を一緒に保持するだけでなく、組織内の細胞が互いに通信することも可能にする。
血液凝固は、細胞コミュニケーションにおける細胞外マトリックスの役割の一例を提供する。 血管を覆う細胞が損傷すると、それらは組織因子と呼ばれるタンパク質受容体を表示します。, 組織因子が細胞外マトリックス中の別の因子と結合すると、血小板が損傷した血管の壁に付着し、隣接する血管平滑筋細胞を刺激して収縮(したがって血管を収縮)し、血小板を刺激して凝固因子を産生する一連のステップを開始する。
細胞間接合
細胞はまた、細胞間接合と呼ばれる直接接触によって互いに通信することができる。 植物と動物の細胞がこれを行う方法にはいくつかの違いがあります。, Plasmodesmata(単数=plasmodesma)は、植物細胞間の接合であり、動物細胞の接触にはタイトおよびギャップ接合、およびデスモソームが含まれる。
一般に、隣接する植物細胞の原形質膜の長い伸びは、各細胞を囲む細胞壁によって分離されているため、互いに触れることができない。 Plasmodesmataは、隣接する植物細胞の細胞壁の間を通過し、その細胞質を接続し、シグナル分子および栄養素を細胞から細胞に輸送することを可能にする多数のチャネルである(図6a)。,
タイトジャンクションは、隣接する二つの動物細胞間の水密シールです(図6b)。 タンパク質は細胞を互いにしっかりと保持します。 この堅い付着は材料が細胞の間で漏ることを防ぐ。 タイトジャンクションは、典型的には、内臓器官および空洞を覆い、皮膚の大部分を構成する上皮組織に見出される。 例えば、膀胱を覆う上皮細胞のタイトな接合部は、尿が細胞外空間に漏れるのを防ぎます。,
また、動物細胞にのみ見られるデスモソームは、隣接する上皮細胞間のスポット溶接のように作用する(図6c)。 彼らは、皮膚、心臓、および筋肉のように、伸びる器官および組織におけるシート状の形成において細胞を一緒に保つ。
動物細胞におけるギャップ接合は、隣接する細胞間のチャネルであり、細胞が通信することを可能にするイオン、栄養素、および他の物質の輸送を可能にするという点で、植物細胞における原形質連鎖のようなものである(図6d)。 しかし、構造的にはギャップ接合と原形質連絡は異なる。,
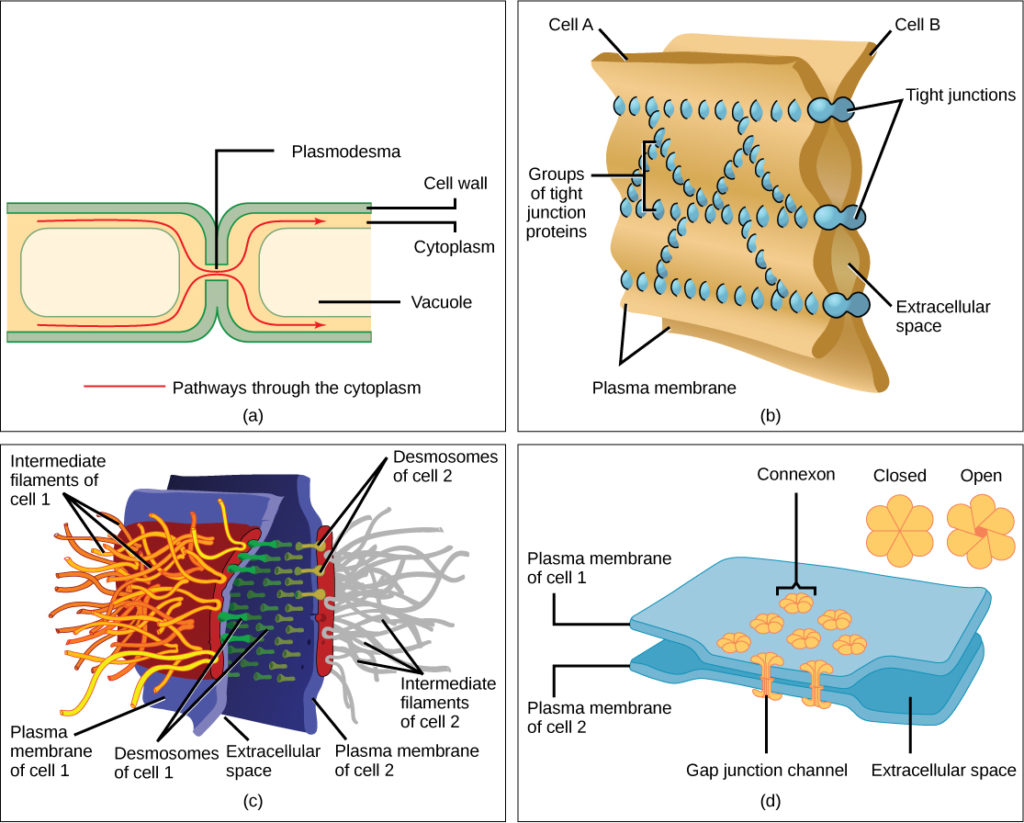
図6. セル間の接続には四種類があります。 (a)plasmodesmaは、隣接する二つの植物細胞の細胞壁間のチャネルである。 (b)タイトジャンクションは、隣接する動物細胞を結合します。 (c)デスモソームは二つの動物細胞を一緒に結合する。 (d)ギャップ接合は、動物細胞間のチャネルとして作用する。 (credit b,c,d:Mariana Ruiz Villarealによる作品の修正)
貢献してください!
このページを改善するより多くを学ぶ